Evolution of parasite life history
Life history theory
In general, life history refers to the
"choices" (not really choices, but evolved strategies) that an
organism makes about the timing of growth, reproduction, and death.
Individual life histories are interesting of themselves, and also
underlie demography and population dynamics, which we will study
later. The microevolutionary struggles of individuals for fitness,
and possibly the selection between lineages for survival, determines
the life-history patterns we see among taxa.
Applications: conservation biologists routinely use life history
and demography to figure out the best ways to conserve species
(sea turtles); to eliminate pest species; and to understand how
species will evolve in response to human harvesting pressure
(marine fisheries). People haven't actually done this for
parasites (mostly they're just trying to find chemical targets
that work, and hope that parasites don't evolve too much in
response), but they could ...
As always in this class, we can look both at the life histories of
parasites themselves, and how they have evolved to fit the constraints
of a parasitic life cycle, and at how the life cycles of hosts are
affected by parasites.
First, however, we'll discuss life history in general.
Classical life history traits are:
- growth rates
- timing of maturity/first reproduction
- size at maturity
- fecundity
- size of offspring
- timing of reproduction: semelparity vs iteroparity
- longevity
- size of offspring, number of offspring
Obviously, these traits are all tied together, by physiological
correlations (e.g. large size is correlated with high fecundity
and longevity) and tradeoffs (fecundity vs longevity,
size at maturity vs timing of maturity, etc.).
The correlations and tradeoffs depend on the detailed physiology
of organisms and on their environment (in a nutrient-rich environment,
potential growth rates will be higher and so the tradeoff between
early maturity and large size at maturity will be weaker). In any
case, organisms will evolve to maximize their fitness in a particular
environment.
|
RVx = mx + |
Ą
ĺ
t = x+1
|
mt Surv(x,t) |
| |
That is, total reproductive value is the current
reproductive output, m, plus (potential) future reproductive output
times the probability of surviving that long.
Tradeoffs come in the tradeoff between current reproduction and future
reproduction and survivorship, or indirectly between current
reproduction and allocation to other traits (e.g. size) that will
change future survivorship and reproduction.
r/K selection
One classical paradigm for how all those different life-history
characters are connected is the r-K tradeoff.
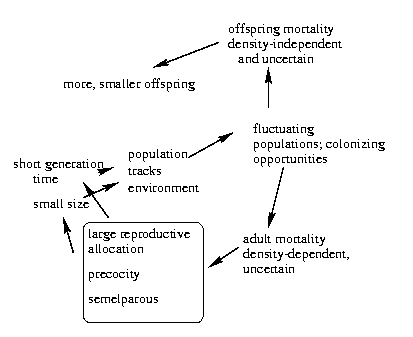 Begon, Harper and Townshend give these connections between
the various traits of (in this case) r-selected organisms,
which are all ultimately driven by the fluctuating nature
of the environment.
Begon, Harper and Townshend give these connections between
the various traits of (in this case) r-selected organisms,
which are all ultimately driven by the fluctuating nature
of the environment.
How would you think about controlling r vs K-selected
organisms?
Parasites are often thought to be purely r-selected.
In particular, it was originally thought that parasites
were unusually small and fecund.
The real story is, of course, more complicated.
Body size
Non-parasitic lineages typically obey Cope's Rule:
lineages increase in body size through evolutionary time.
(Q: why?)
(But there is a tendency for
species body-size distributions to be right-skewed
(Fowler and MacMahon 1982, Blackburn and Gaston 1994a):
how does this happen? Suggestion: within-lineage evolution
for larger size, but between-lineage selection through
higher speciation rates in smaller animals (smaller =
quicker life cycle = faster evolutionary rates?) and
possibly more evolutionary inertia (ditto).)
Do parasites obey Cope's Rule?
The typical assumption is that parasites evolve to be
smaller, but as Poulin's discussion suggests
this is really caused by a confusion between the fact
that parasites are small (they have to be, so they
can live in or on their hosts) and the possibility that
they have gotten smaller. Small size is a necessary
pre-adaptation for parasitism (as long as we are sticking
to the "close association" part of the definition of parasitism):
we wouldn't expect to see elephants becoming a parasitic lineage ...
Changes in body size
Poulin looks at the body-size distributions of
various families of parasites. He is comparing within
families, which takes care of part of the problem
with phylogenetic contrasts, but does not correct (e.g.) for the
possibility that smaller species speciate more rapidly.
Also, there is probably detection bias: smaller species
are harder to discover.
Nevertheless, just looking at body-size distributions
for nematodes, copepods, and isopods, shows that free-living
stages are smaller than or about the same size as parasites of
invertebrates, and typically smaller than parasites of vertebrates.
We have to be careful interpreting these pictures:
- copepods: a careful phylogenetically controlled analysis of the data
shows that body sizes are statistically the same in free-living
and vertebrate-parasite species (9 comparisons),
and larger in fish parasites (9
comparisons).
- isopods: phylogenetic contrasts actually suggest
that isopods have become smaller in parasitic
lineages (despite the pattern of body-size distributions).
- amphipods: become smaller (although not much).
(However, note that these symbiotic amphipods are
mutalists: how might that change the story??
Ecological correlates of body size
- Primarily host body size; this very often comes
up as significantly correlated with parasite body
size, especially after correcting for phylogeny etc.
- in ectoparasites (but not endoparasites), environmental
temperature/fluctuation etc. seems to matter:
colder=larger
- sexual dimorphism: often (e.g. copepods)
females increase in size while males remain the
same size (strongly points to the importance
of the size/fecundity/female lifespan connections,
which we will emphasize again in a moment)
- host longevity??
Fecundity
- Comparisons with free-living stages:
interesting idea, but practically no data (!)
- within parasite lineages:
correlation with host body size? (Yes, see below.)
- Correlation with life cycle type? (No.)
- Tradeoff in many taxa (but not nematodes)
between egg size and number (fairly obvious
physiological mechanism: the direction of
the tradeoff depends on life-history details,
in particular what environment the offspring
will face on hatching and whether size will
provide an advantage in survival or competition
(e.g. latitudinal gradients, poikilothermic
vs homeothermic hosts)
Correlations and suites of characters
Skorping et al 1991:
- small/fast/low fecundity/short reproduction
(trichostrongyles)
to large/long development/high fecundity (ascarids)
- explanations:
lack of tradeoffs within host environment;
independence of offspring success on initial
size (although cf Loker); fixed windows
(host generation time, transmission windows)
Morand 1996:
- a bit confusing: corrects for body size and
for phylogenetic contrasts, but not both
at once
- allometry between size and fecundity: fecundity increases
with size (slower than linearly)
- age at maturity linked with size at maturity
- no connection between fecundity and mortality!
- not that much difference between free-living and vert.
parasitic groups
- mortality appears to drive life history,
but not necessarily in a simple "r-selected" way
Host life-history responses to parasites
Just as a final reminder, there's a strong possibility that parasites
can affect the life histories of their hosts.
Here's the abstract from a recent study of mosquitoes infected
by microsporidian parasites:
Interactions between host and parasite life history traits : we showed
that females of C. pipiens increase their developmental rate and thus
pupate earlier when infected by the mirosporidian parasite
V. culicis. This decrease of the developmental time, in agreement with
theoretical predictions, comes at the cost of reduced adult size and
hence reduced fecundity (Agnew et al. 1999). Because the reaction is
manifested by the infected individuals themselves, this life-history
adjustment reflects phenotypic plasticity.
Agnew, P., Bedhomme, S., Haussy, C. and Michalakis, Y. 1999. Age and size at maturity of the mosquito Culex pipiens infected by
the microsporidian parasite Vavraia culicis. Proc. R. Soc. Lond. B (in press).
What does the existence of plasticity say about the evolutionary
history and ecological prevalence of the mosquito-microsporidian association?