Parasite life cycles: evolution and consequences
Where and why do complex life cycles come from?
Parasite life cycles: they're cool, and seemingly improbable,
natural history ... many parasites (as we will see in a bit, after the
inevitable digression) have what seem to be ridiculously
baroque ways of producing the next generation of offspring,
involving transfers between multiple hosts.
Complex life cycles involve shifts between hosts, but
can also involve a succession of physiologically or
morphologically different stages within a single
host (e.g. using different tissues). There is a close
parallel between complex life cycles in parasites and
metamorphosis, e.g. in amphibians or
lepidoptera. In each case, organisms undergo
major physiological/morphological rearrangement as
part of their life cycle.
Definitions and examples
Many parasites (especially "classical" metazoan parasites)
go through multiple species in their lifetimes.
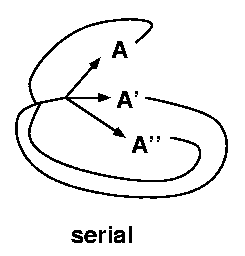
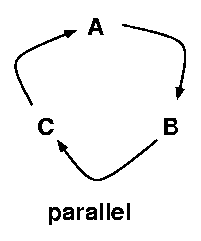
Life cycle complexity is the use of multiple host species in
serial (all host species must be used in
sequence to complete a single parasite life cycle),
while specificity is the use of multiple host species
in parallel (individual parasites can pick
one of several hosts to complete a particular life
cycle stage).
- Parasites that use only a single host species in their
life are called
monoxenic;
- those with multiple hosts are
called heteroxenic.
- Definitive hosts are those in which sexual reproduction takes
place (there is some reason to argue that definitive hosts are
special, since sexual reproduction
can only occur once in the life cycle, but a parasite could
shift its definitive stage from one host to another);
- intermediate hosts are all the others.
- Paratenic hosts are hosts that are used only for transport;
no life cycle stages are completed within the host.
- Vectors are hosts that facilitate
transport from one host to another, although what is a
"vector" and what is not depends on your focus.
To a mosquito, a human might serve as a vector
for malaria to get from one mosquito to another!
The malaria parasite provides an example of a complex life
cycle with two hosts and multiple life stages within each
host. Combes (p. 590) provides a good description and
picture of the sequence from sporozoites, hepatocytes (human liver),
merozoites (human red blood cells), gametocytes (mosquito guts),
ookinetes (travel to mosquito salivary glands), back to sporozoites.
Sexual reproduction takes place in the mosquito, which is therefore
the definitive host. Asexual reproduction occurs at several
points around the cycle, within both humans and mosquitos.
Other examples of transmission include:
- Direct transmission:
most viruses, bacteria;
many plant fungi;
many intestinal nematodes
- Two hosts/one intermediate host: e.g. Schistosoma mansoni
(trematode), causes human schistosomiasis:
human (or rodents, cattle, baboons, dogs) to snail.
By some definitions, this could also include classical
vector-borne parasites: malaria (protozoan),
filarial worms
- multiple intermediate hosts:
e.g. Paragonimus (another trematode):
mammals to snails to crayfish.
Alaria spp.: wild carnivores,
snails, tadpoles, mice/rats/snakes (paratenic)
Combes (p. 38) mentions Halipegus ovocaudatus,
a trematode
which has a four-host cycle from amphibian to mollusk
to crustacean to insect.
Q: is there a limitation on the
number of hosts in a life cycle? Why doesn't it keep increasing? Is
this obvious, or not?
Transition methods:
- passive movement followed by uptake of eggs/larvae by feeding,
inhalation, etc.
- active locomotion (by eggs/larvae/parent),
followed by penetration of the host
and possible migration within the host
- next host feeds on the previous host (predation or
incidental: behaviorally mediated or not).
It makes sense that trophic transmission
is common, since trophic interactions are probably
the most common way that individuals of different
species come into close physical contact with
each other in the wild.
Why??
Complex life cycles seem hard to justify on the face of it.
Why would an organism employ such a dicey strategy?
This question has led to creationist
arguments, but for now let's stick to the evolutionary ones ...
We come up with the
same classes of answers that always come up in
response to evolutionary "why" questions:
adaptationist and drift/neutral arguments.
- neutral arguments say that organisms found themselves
switching by hosts essentially by accident (e.g. when the
host they were in was swallowed by an individual of another
species). Once a lineage of parasites was stuck in this
cycle, they could lose the ability to have a shorter cycle
via mutation and drift.
- there are several kinds of adaptationist arguments:
- organisms that find themselves in other hosts (as in
the example above) could experience tradeoffs
that encouraged the evolution of obligately complex
life cycles
- adding another host could actually be advantageous,
providing transport between hosts or allowing parasites
to limit their virulence within different life stages;
- Smith Trail has suggested "host suicide" as a host
adaptation (committing suicide to lessen the risks of
parasitism for one's kin) that could then be perverted
and turned to advantage by parasites.
How?
Testing these hypotheses also brings us back to a familiar refrain.
There are essentially four ways of testing hypotheses on parasite
evolution:
- Broad comparative: use a large database to try to
look at ecological and evolutionary correlates of the trait
in question (in this case, life cycle complexity) to see whether
the patterns that emerge either support available hypotheses
or suggest new ones;
- Narrow comparative: test a closely
related set of species in similar environments that show
some interesting difference in parasitism, and try to
infer the reasons for their differences;
- Experimental: manipulate organisms
in the field or in the laboratory;
- Phylogenetic: test the relationship
between the phylogeny of some group and the pattern
of changes in parasitism.
For this particular case, we have examples of two of
the above:
- Combes discusses two examples (pp. 159-163) of "narrow"
comparative approaches to life cycle complexity.
I briefly discussed his example of Bothriocephalus, where
there are two closely related parasites (gregarius
and barbatus) in similar ecological situations
(on turbot and brill in similar environments) that behave
differently (barbatus has a 2-host cycle,
gregarius has a 3-host cycle including gobies
as a paratenic host). This supports one of the adaptive
arguments above.
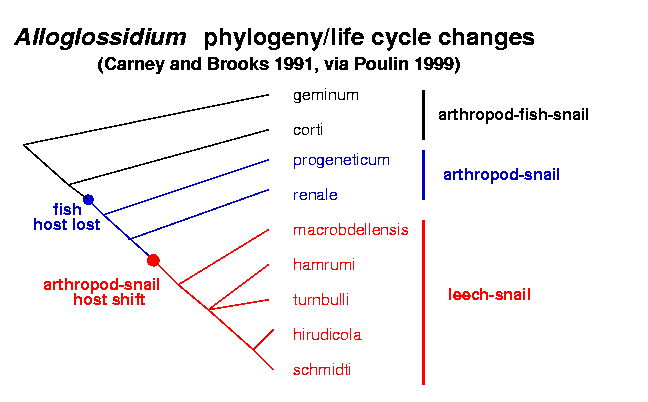
- Combes (p. 165) discusses the evolution of
shorter life cycles in Alloglossidium;
the following phylogeny
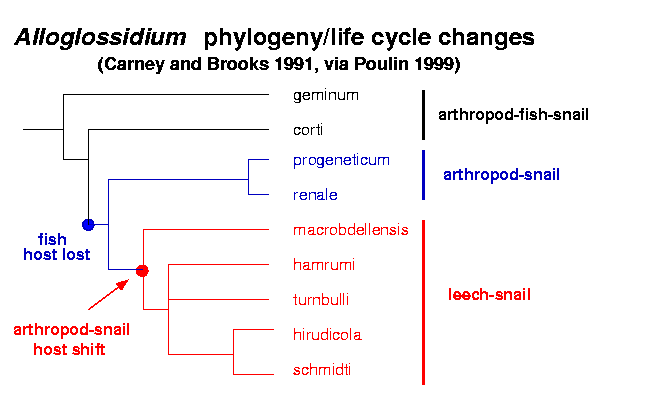 suggests that a 3-host cycle is ancestral.
(Note that drawing the same phylogeny
this way
would be equivalent.)
Of course,
this doesn't tell us why it happens, just
that it happens. Combes waves his hands a bit about
possible reasons for the shortening in this and other systems.
Q: how could you go about testing/establishing
hypotheses for why this had happened?
suggests that a 3-host cycle is ancestral.
(Note that drawing the same phylogeny
this way
would be equivalent.)
Of course,
this doesn't tell us why it happens, just
that it happens. Combes waves his hands a bit about
possible reasons for the shortening in this and other systems.
Q: how could you go about testing/establishing
hypotheses for why this had happened?
Background from Morand et al. 1995
{kind=link}